2 分子生物学实验技术
2.1分子克隆及相关实验技术
2.1 分子克隆及相关实验技术
2.1.1分子克隆
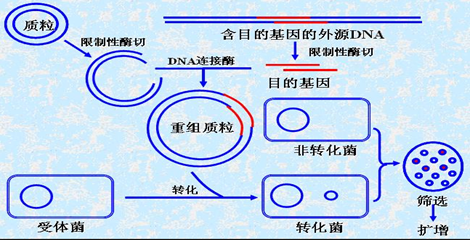
简介(INTRODUCTION)
完整的分子克隆可以简述为“分→切→接→转→筛→表”六个实验,如上图。PCR、酶切、连接、转化是传统基因克隆方法获得重组质粒必不可少的步骤。用PCR方法扩增要克隆的目的基因,限制性内切酶使插入片段和克隆载体之间建立互补配对的碱基序列,再经连接酶的连接反应完成插入片段和线性载体之间的重组。考虑到重组效率,通常要先将连接的产物转化到用于扩增质粒的克隆大肠杆菌感受态细胞中,进一步筛选正确的重组子。
PCR:即多聚酶链式反应。它是一种用于放大扩增特定的DNA片段的分子生物学技术,它可看作是生物体外的特殊DNA复制,PCR的最大特点是能将微量的DNA大幅增加。反应过程分为三步:变性,退火,延伸。变性:双链DNA解旋形成单链。退火:特定引物与DNA单链模板结合。延伸:从5’端向3’端延伸。
酶切:限制性内切酶能特异性地结合于一段被称为限制性酶识别序列的DNA序列之内或其附近的特异性位点上,并切割双链DNA。分子克隆实验中最常用的是Ⅱ型酶,且是那些识别4个或6个碱基对的限制性内切酶。Ⅱ型限制性内切酶的识别顺序是一个回文对称顺序。切割后得到的是带粘性末端或平末端的线性DNA。
连接:DNA连接酶催化双链DNA分子中相邻碱基的5’-P末端与3’-OH间形成3’,5’-磷酸二酯键。一个DNA片段的5’-P末端与另一个3’-OH末端相靠时互近,在DNA连接酶的作用下,有Mg2+,ATP存在的缓冲系统中可以被连接起来而形成重组分子。常用的DNA连接酶是T4 DNA连接酶,其作用底物是双链的DNA分子或RNA:DNA杂交分子,可以连接粘性末端、平末端。
转化:体外连接的DNA分子必须尽快引入寄主细胞,否则会很快降解,而且只有导入到细胞中后,才能扩增及研究其功能的表达,细胞转化是常用也是最有效的方法之一,细菌转化是指裸露的(或纯化的)DNA被细菌吸收而导致基因转移的现象。大肠杆菌的感受态需诱导。将对数生长期的细菌放入0 °C的CaCl2低渗溶液中,细胞膨胀,形成原生质球,诱导感受态;DNA加入后,形成抗DNase的羟基-磷酸钙复合物,粘附于原生质球表面,42 °C热冲击,DNA被吸收,将细胞混合物涂布于合适的(筛选)培养基培养几小时,细胞得以恢复和增殖,转化基因得以表达,然后根据合适的选择条件可以区分转化细胞和非转化细胞。
材料(MATERIALS)
r PCR试剂
核酸模板、引物(10 μM,上下游引物)、dNTPs Mix、MgCl2、DNA聚合酶、HF或GC buffer、DMSO、ddH2O
r 酶切试剂
限制性核酸酶切酶比如Sal I HF、Not I HF、酶切buffer 4、insert or vector
r 连接试剂
T4 DNA连接酶、10×T4 DNA连接酶buffer
r 转化试剂
感受态细胞(如DH5α、TOP10)、抗性LB平板、无菌无抗的SOC或LB培养基
实验步骤(PROCEDURE)
PCR部分:
1.建立PCR反应体系(以Pfu-XI DNA聚合酶为例)
试剂 | 20 μL | 50 μL |
5×HF/GC Buffer | 4 μL | 10 μL |
10×dNTP (2.5 mM each) | 2 μL | 5 μL |
50 mM MgCl2 | 0-0.2 μL | 0.5 μL |
Pfu-XI polymerase | 0.2 μL | 0.2-0.5 μL |
Primers | 0.5 μL+0.5 μL | 1 μL+1 μL |
DNA template | 0.1-1 μL | 0.1-1 μL |
DMSO*(一般不加) ddH2O | 0.2 μL Add to 20 μL | 0.5 μL Add to 50 μL |
选择薄壁的PCR管,按照上述体积系数,将上述物质依次加入混匀。
▲关键步骤:
酶催化反应通常最后加入酶,并冰上操作;DMSO能够减少模版二级结构,从而促进引物与模板的结合,但也会增加突变的概率;在执行PCR扩增程序时,最好每管20~50 μL,过大的体积会使PCR仪中的反应溶液的温度变化的均一性降低,反应条件不准确;具体情况具体分析。
2.建立PCR反应程序(以Pfu DNA聚合酶为例)
Pfu-XI DNA聚合酶常用的PCR扩增程序
预变性 | 98 °C 1~5 min | |
28~35个循环 | 变性 | 98 °C 10 s |
退火 | 62 °C 10 s | |
延伸 | 72 °C 1 kb/15~30 s | |
再延伸 | 72 °C 5 min | |
保温 | 12 °C 10 min | |
▲关键步骤:
♫不同的DNA聚合酶有不同的效率:Pfu-XI:1 kb/20 sec,Taq:1 kb/1 min;
♫ 若是以菌落或菌液为模板,预变性时间可适当提高至3-5 min;
3.琼脂糖凝胶电泳实验鉴定 PCR 结果
▲选择合适的琼脂糖凝胶浓度及合适大小的DNA Marker。
4.PCR 产物纯化回收
若条带产物较纯,推荐用PCR产物回收试剂盒回收,若条带较杂可用胶回收。操作步骤见试剂盒说明书。
酶切部分:
1.建立酶切体系:
2.插入片段的酶切体系:插入片段可以是已经纯化回收的 PCR 产物或含有插入片段的质粒。以Sal I /Not I 系列的内切酶为例,酶切 PCR 产物的体系如下:
PCR products | 1~2 μg |
10× buffer 3 | 10 μL |
Sal I | 0.5 μL |
Not I H2O | 0.5 μL Add to 50 μL |
放置于37 °C水浴锅,反应2~3 h。
▲注意事项
♫ By definition, 1 U restriction enzyme can cleave 1 μg DNA plasmid in 1 hour. Sal I, Sal I HF, Not I HF is 20 U/μL, which means in most cases, 0.5 μL or 10 U is more than enough.
♫ 选择合适的内切酶和该酶所对应的特定的酶切buffer,每种内切酶都有其高效反应buffer。如果不清楚双酶切的共用buffer时,建议去NEB官网查下具体的酶切信息http://nebbuffer;
♫ 有些内切酶的反应可能要求加入BSA(胎牛血清中的一种球蛋白:牛血清白蛋白,是酶的稳定剂,防止酶的分解和非特异性吸附),是否添加可在NEB官网查到;
♫ 严格控制酶切体系中的甘油浓度在10%以下,否则容易出现酶切的“星活性”现象,即酶切的专一性被放宽,酶切“序列识别”的严谨度下降(比如从严格识别的6个碱基才能酶切到只要识别4个相似碱基时就可以切割),不但可以切断特异性的识别位点,还可以切断非特异性的位点,出现乱切现象。
♫ 乱切原因:a.低盐浓度(<25 mM);b.高pH值(>pH 8.0);c.存在有机溶剂(如DMSO、乙醇、乙烯乙二醇、二甲基乙酰胺、二甲基甲酰胺、sulphalane等);d.用其他二价离子替代镁离子(如Mn,Cu,Co,Zn 等)。
♫ 乱切解决办法:a. 尽量用较少的酶进行完全消化反应。这样可以避免过度消化以及过高的甘油浓度;b.尽量避免有机溶剂(如制备DNA时引入的乙醇)的污染;c.将离子浓度提高到100~150 mM (若酶活性不受离子强度影响)。
3. 酶切产物回收
酶切产物试剂盒回收法或琼脂糖凝胶胶回收方法
4.克隆载体酶切效果检测(可选步骤):酶切后的载体和完整载体分别转化,对比转化结果。
连接部分:
1. 建立连接反应体系:典型的体系为10 μL反应体系(以T4 ligase & 10×ligation buffer为例)
T4 ligase 10 μL反应体系 |
|
Insert |
|
Vector |
|
10×ligation buffer | 1 μL |
T4 ligase H2O | 1 μL Add to 10 μL |
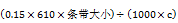
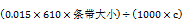
冰上操作,将上述物质加到PCR管中混匀,放置16 °C或室温反应过夜(8~12 h)。为减少蒸发,建议使用PCR管。
▲关键步骤:
载体酶切的质量是连接成功的关键。连接前要确保载体的酶切效果和浓度;连接体系以小体积建立,以增加分子碰撞的机会;合适的insert :vector比值对连接反应的成功贡献巨大。体积比例的设置依据各物质量比(和insert的大小和浓度相关),insert不低于0.3 pmol。体积比值一般选择在1:1到1:8之间,增加insert的量主要是减少载体自连。NEB计算连接反应物质浓度:http://nebiocalculator.neb.com/#!/dsdnaamt
转化部分:
1.从-80 °C取出感受态细胞放在冰上溶解(大约5 min)。做好标记,建议同时取一管感受态细胞做空白对照;
2.从4 °C冰箱取出抗性平板和SOC(根据连接的载体的抗性选择抗性筛选的LB平板),倒置平放在37 °C恒温培养箱中孵育备用。设置水浴锅温度为42 °C;
3.待感受态细胞的最后一粒冰融化时,取连接反应产物的一半(5 μL)转到50 μL的感受态细胞中,指肚轻弹几下EP管底部混匀。冰上放置20~30 min;
4. 轻轻取出转化管固定在浮板上,42 °C热激45~90 s。将EP管的1/2或2/3没于水中,定好闹钟。(60 s通常是理想的热激时间,热激时间取决于所使用的感受态细胞);
5.冰上放置2~10 min,再室温放置2 min;
6.超净台中取500 μL无抗无菌的LB or SOC培养基,37 °C摇床摇60 min复苏细胞(将EP管倾斜45度放置,注意时间不要超过三个小时,此时杂菌可能成为优势菌);
7.4000 rpm离心5 min,取出37 °C孵育的抗性平板并做好标记(质粒名称、抗性、感受态细胞名称、日期、姓名),在无菌操作台中倒掉离心后的上清至100−200 μL,重悬细胞,全部涂布到平板中(涂棒在使用前后在酒精灯外焰干烧一会儿以达到无菌,无菌玻璃珠用前用后要专门分开放置);
8.平板先正置放置在37 °C恒温培养箱放置10 min,使菌液凝在培养基上(可选步骤);
9.将平板倒置放置在37 °C恒温培养箱。培养16 h(一般14~18个小时都正常,24小时后就会杂菌蔓延)。
▲关键步骤:
♫ 转化最好做阴性(如感受态细胞、无抗培养基)或和“假阳性”(如空载体)对照,以确保转化操作是work的,试剂也是可行的,如感受态、SOC/LB、抗性平板等;
♫ 对于特别大的质粒可以用电转化的方法,但一般很少用到,此处不作介绍;
♫ 因为转化后的培养时间较长,建议转化操作选在一天的下午进行,这样便于第二天上午观察检测结果。
2.1.2 引物设计
简介(INTRODUCTION)
引物设计是一小段单链DNA或RNA,在核酸合成反应时,作为每个多核苷酸链进行延伸出发点而起作用的多核苷酸链。通常指的设计引物都是在已知模板序列的情况下进行。以目的基因为模板设计一对特异寡核苷酸引物,从而进行DNA的体外合成反应。在引物的3′-OH上,核苷酸以二酯链形式进行合成,因此引物的3′-OH,必须是游离的。PCR引物设计目的是在扩增特异性和扩增效率两个目标上取得平衡。
基本原则(PRINCIPLE)
1. 引物长度
一般引物长度为18~30碱基。确保退火温度不低于54 °C的最短的引物可获得最好的效率和特异性。每增加一个核苷酸引物特异性提高4倍,这样,大多数应用的最短引物长度为18个核苷酸。引物长度的上限并不很重要,主要与反应效率有关。由于熵的原因,引物越长,它退火结合到靶DNA上形成供DNA聚合酶结合的稳定双链模板的速率越小。另外引物太长,PCR的最适延伸温度会超过聚合酶的最佳作用温度(70 °C),从而降低产物的特异性。
2. GC含量
一般引物序列中G+C含量一般为40%~60%,一对引物的GC含量和Tm值应该协调。若是引物存在严重的GC倾向或AT倾向则可以在引物5’端加适量的A、T或G、C尾巴,并且相应的减少或增加引物的长度。
3. 退火温度(Tm值)
一定盐浓度条件下,50 °C寡核苷酸双链解链的温度。Tm值计算网站链接见附录。PCR扩增中的复性温度一般是较低Tm值引物的Tm值减去5~10 °C。如果引物碱基数较少,可以适当提高退火温度,这样可以使PCR的特异性增加;如果碱基数较多,那么可以适当减低退火温度,使DNA双链结合。一对引物的退火温度相差4~6 °C不会影响PCR的产率,但是理想情况下一对引物的退火温度是一样的,可以在55~75 °C间变化。
4. 模板的二级结构区域
对于RT-PCR或Q-RT-PCR的引物设计,选择扩增片段时最好避开模板的二级结构区域。用有关计算机软件可以预测估计目的片段的稳定二级结构,有助于选择模板。但是对于蛋白的克隆表达,有时候必须要选择到二级结构区域作为模板,故酌情选择。
5. 引物的位置(主要针对RT-PCR或Q-RT-PCR检测引物的设计
引物的序列应位于基因组DNA的高度保守区,且与非扩增区无同源序列,这样可减少引物与基因组的非特异结合,提高反应的特异性。若以cDNA为模板,则首先应尽量使引物和产物保持在mRNA的编码区域内;其次,尽量把引物放在不同的外显子上,以便使特异的PCR产物与从污染DNA中产生的产物在大小上相区别。
6. 引物自身的检查表达
1) 引物的二级结构:引物自身不应存在互补序列,否则引物自身会折叠成发夹状结构,特别是引物的末端应避免回文结构,这种二级结构会因空间位阻而影响引物与模板的复性结合。若用人工判断,引物自身连续互补碱基不能大于3 bp。两引物之间不应该存在互补性,尤应避免3’端的互补重叠以防引物二聚体的形成。一般情况下,一对引物间不应多于4个连续碱基的同源性或互补性。
2) 上下游引物的互补性:一个引物的3’末端序列不允许结合到另一个引物的任何位点上。
3) 3’末端:引物3’端是延伸开始的地方,因此要防止错配就从这里开始。3’端不应超过3个连续的G或C,因这样会使引物在G+C富集序列区错误引发。3’端也不能形成任何二级结构和不能发生错配。如扩增编码区域,引物3’端不要终止于密码子的第3位,因密码子的第3位易发生简并,会影响扩增特异性与效率。如果可能的话,每个引物的3’末端碱基应为G或C。
4) 5’端对扩增特异性影响不大,因此,可以被修饰而不影响扩增的特异性。引物5’端修饰包括:加酶切位点;标记生物素、荧光、地高辛、Eu3+等;引入蛋白质结合 DNA 序列;引入突变位点、插入与缺失突变序列和引入一启动子序列等。额外的碱基或多或少会影响扩增的效率,还加大引物二聚体形成的几率,但是为了下一步的操作就要作出适当的“牺牲”。
实验步骤(PROCEDURE)
1. 在NCBI(PUBMED)上搜索目的基因,找到该基因的mRNA序列,在CDS选项中,找到编码区所在位置,复制到Seq Builder建立序列文件。
对于RT-PCR和Q-RT-PCR所需的鉴定引物,建议用Primer Premier5搜索最佳打分引物。即Tm值在55~70 °C之间,GC%含量在45~55%之间,上下游引物Tm相差2 °C以下,并且不存在发卡、二聚体、引物间交叉二聚体和错配等二级结构。
对于蛋白的克隆表达所需引物:Full-length蛋白的克隆,老老实实按照ATG…TAA起始密码子和终于密码子全部扩增出来。
2. 设计PCR引物时需要在酶切位点序列的外侧添加保护碱基以提高酶切时的活性。实验室常用保护碱基为:GTC、CTA
针对性建议(TROUBLESHOOTING)
1. 引物纯化主要方式:无特殊实验要求,克隆实验一般推荐RPC纯化方式。
2. 一般默认模板序列排列方式是5′-3′为主链,因此上游引物与模板是一致的,而下游引物是与模板序列反向互补的序列,在复制设计好的引物时切忌检查。本实验常用软件Seq Builder里,标记引物使用filled arrow,方便于将设计好的引物直接复制粘贴。
3. 设计的引物太短,特异性降低;引物太长,影响产物的生成。建议结合引物序列GC含量和Tm值,选择最适的长度。值得注意的是引物的长度是指与模板DNA序列互补的部分,不包括为后续克隆而添加的酶切位点以及额外保护碱基序列。
4. 引物的3’末端的第一和第二个碱基影响DNA聚合酶的延伸效率,故其影响PCR反应的扩增效率和特异性。建议引物3’末端选择G或C,因为他们形成的碱基配对比较稳定。
5. 引物使用浓度:建议0.1~1.0 μM,默认是1 μM。浓度太高会导致错配、引物二聚体增加;浓度太低导致PCR效率降低。
简介(INTRODUCTION)
Gibson assembly是一种one step,one pot的快速基因组装方法。它只需要将基因片段和需要的三种酶混合在同一个管内在50 °C下培养15~60 min就可以得到组装好的DNA。
|
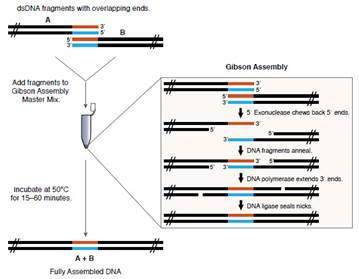
装配原理基于DNA片段间的重叠区域,过程依赖于三种酶:DNA外切酶(T5 exonuclease),高保真DNA聚合酶(Phusion polymerase)和耐热DNA连接酶(Taq DNA ligase)的共同作用。首先,T5核酸外切酶消化DNA片段的方向是从5’端到3’端。每个DNA片段分别形成一个单链的突出部分,由于这两个相邻的突出片段有一部分具有同源性能够互补,所以DNA片段退火后,互补的序列重新配对连接。然后,在空缺的部分DNA聚合酶以另一条DNA单链为模板,沿3’方向将对应的脱氧核苷酸连接到单链上,填补缺口。最后,连接酶将两条DNA单链黏合起来,密封裂缝。这样具有重叠区域的DNA片段就组装成一整条DNA分子了。最近我们实验室所使用的是5×T5试剂(里面只有一种T5 exonuclease外切酶,没有其他组分),依靠细菌自身的重组能力进行重组转化克隆。下面是组装的示意图。最近文献和我们的结果显示,只采用T5 exonuclease一种酶,等DNA退火后,可以直接转化入感受态细胞,利用细胞内DNA修复机制修复,从而复制完整双链质粒。这种方案效率更好,但是不能用电转法。
材料(MATERIALS)
r 试剂(REAGENTS)
NAD,H2O,1 M MgCl2,1 M DTT,10 mM dNTP mix,1 M Tris-HCl pH 7.5,50% PEG-8000,2.7 mg/mL Tag ligase,0.85 μg/mL T5_ExO,Phusion(1×)、LB培养基、抗生素(据载体而定)、LB平板(相应抗性)
r 实验前准备(SETUP)
于冰水浴中配制如下反应体系。如果不慎将液体粘在管壁,可通过短暂离心使其沉入管底。
4x isothermal assembly buffer | 加入量 |
NAD | 20 mg |
H2O | 300 μL |
1 M MgCl2 | 300 μL |
1 M DTT | 300 μL |
10 mM dNTP mix | 600 μL |
1 M Tris-HCl pH 7.5 | 3 mL |
50% PEG-8000 | 3 mL |
加水到7.5 mL,每管分500 μL存于-20 °C或-80 °C冰箱,够3000个反应。
2× assembly mix: best combination: | 加入量 |
|
100 reaction | 1 mL |
|
2.7 mg/mL Tag ligase | 10 μL |
|
0.85 μg/ mL T5_ExO | 100 μL | |
Phusion(1×) | 25 μL |
|
4× isothermal assemble buffer | 500 μL |
|
加水365 μL,存于-20 °C冰箱,有效期1年。
实验步骤(PROCEDURE)
1. 设计引物通过PCR的方法在DNA片段的两端加上同源片段,NEB推荐同源片段的长度为15-40 bp,同时要求这部分对应的退火温度高于50 °C。
2. 进行DNA纯化:PCR产物进行琼脂糖电泳,胶回收,或者PCR产物回收试剂盒回收。
3. 进行重组反应,以20 μL反应体系为例:
组分 | 加入量 |
Isothermal assembly Master Mix | 10 μL |
线性化载体 | 10~100 ng(1~2 μL) |
插入片段 | 8~9 μL (3~5倍载体) |
Sterilized ddH2O | 补足至20 μL |
50 °C反应15~60 min。
4. 转化10 μL的重组反应产物,涂板及克隆鉴定同传统分子克隆方法。
针对性建议
1. 线性化载体的制备有两种,第一种是使用内切酶对克隆载体进行酶切,此方法的优点是载体的突变率较低,高保真,但缺点是酶切不完全会使假阳性克隆空载体的出现影响单克隆的鉴定。第二种是反向PCR扩增制备线性化克隆载体,再采用胶回收或者Dpn I消化以去除环状载体质粒模板的影响,此方法的优点是假阳性克隆较少,但缺点是因PCR扩增的条带较长,突变的概率也会相应增加。
2. 在进行DNA纯化时,PCR产物最好进行琼脂糖电泳,然后进行胶回收以去除PCR扩增时加入环状质粒模板对克隆鉴定的影响。
3. 在选择克隆位点时,应避免选择克隆位点上下游50 bp内有重复序列的区域。当克隆位点上下游20 bp区域内GC含量均在40%~60%范围之内时,重组效率将达到最大。如这部分区域GC含量高于70%或者低于30%,重组效率会受到较大影响。
4. Gibson组装克隆法缺点之一是适用与长度超过200 bp的片段的组装;缺点之二是如果黏性末端形成稳定的二级结构,如发夹结构或者茎环结构,那么成功率会大受影响,优点是可以同时进行多个片段的组装克隆。
5. 在进行重组反应时,载体用量一般在50~100 ng较好,载体和片段的摩尔比为1:1至1:3,当片段小于200 bp时,片段的用量可增加到载体的5倍量。
6. 50 °C反应的时间最好不要超过60分钟。
2.1.4 质粒抽提-大抽
本方法使用QIAGEN(德国凯杰)质粒提取试剂盒。QIAGEN-tips内独特的阴离子交换树脂专为纯化核酸设计。它独特的分离特性,使纯化的DNA的纯度相当于连续2次CsCl梯度离心得到的DNA纯度。预装的QIAGEN-tips利用重力流原理工作,缩短了纯化质粒的手动操作时间。整套QIAGEN质粒纯化体系避免使用苯酚、氯仿、溴化乙锭和CsCl等有毒物质,减小对使用者和环境的危害。
应用:QIAGEN Plasmid Kits纯化得到的质粒DNA适合多种应用,如:转染、克隆、PCR、体外转录。
实验步骤(PROCEDURE)
1. 细菌培养物的生长
挑取单菌落接种到2~5 mL含有适当抗生素的LB液体培养基,37 °C,200 rpm/h的摇床中振荡8小时。用LB培养基以1/500~1/1000的比例稀释含菌落的培养基。高拷贝质粒用100~200 μL含菌落液体培养基接种到100 mL LB培养基中;低拷贝质粒用250~500 μL含菌落培养基接种到250~1000 mL LB培养基,37 °C,200 rpm/h的摇床中振荡12~16小时。
2.细菌的收获和裂解
3000 g,4 °C离心15 min收获细菌。加入20 mL Buffer P1重悬细菌,后分到两个50 mL离心管。再各添加10 mL Buffer P2,剧烈晃动离心管4~6次使其彻底混匀,室温放置5 min裂解。然后各加入10 mL Buffer P3到此溶菌产物中,晃动离心管4~6次混匀。冰育15~20 min,中和碱。4000 rpm,4 °C离心30 min。将上清液倒入针口密封的过滤器中,打开针口的密封盖,轻轻挤压活塞将上清液过滤到一新的50 mL离心管中。再一次去除细胞渣滓。
3.质粒DNA的纯化
1) 将QIAGEN-tip 500的柱子挂在一个接废液容器上,加10 mL Buffer QBT,让管中溶液滴下排空,润洗。
2) 将第6步上清液加入QIAGEN-tip柱子中让其透过树脂慢慢滴下,用废液容器接废液。
3) 用2×30 mL Buffer QC 洗QIAGEN-tip柱。
4) 再用15 mL Buffer QF 洗脱DNA,用离心管收集DNA洗脱液洗脱。
5) 加入10.5 mL异丙醇到洗脱液中使DNA沉淀下来,混匀。6000 g,4 °C用尖底管离心30 min,离心后小心倒出上清液,倒扣管子。
6) 37 °C烘干离心管底部的质粒5~10 min;用500 μL无内毒素的Buffer EB(小抽Kit中有)重新溶解DNA。
2.1.5 化学法E. Coli感受态制备
简介(INTRODUCTION)
在自然条件下,很多质粒都可通过细菌接合作用转移到新的宿主内,但在人工构建重组质粒过程中,由于缺乏转移所必需的mob基因,重组质粒一般不能自主进入受体菌,而是需要帮助才能穿过细胞膜进入细胞内,随着受体菌的生长、增殖,重组质粒才得以复制、扩增。
1972年Cohen等人采用CaCl2处理大肠埃希菌细胞,发现其能作为一个很好的受体有效接受外源质粒DNA。CaCl2制备感受态细胞的方法具有操作简单、重复性好、转化率高等优点。在这种化学方法中,低温和钙离子的主要作用是破坏细胞膜上的脂质阵列,细菌在CaCl2低渗溶液中膨胀为球形,转化混合物中的DNA形成羟基-钙磷酸复合物黏附于细胞表面,经42 °C热休克后促使细胞吸收外源DNA。在丰富培养基中生长后,细胞复原并分裂增殖,重组质粒中的基因在被转化的细菌中得到表达,在选择性培养基平板上即可筛选所需要的转化子。
除了CaCl2法之外,还有RbCl(KCl)法制备感受态细胞,RbCl(KCl)法制备的感受态细胞转化效率较高,我们实验室常用TB法制备感受态细胞,这种方法是在CaCl2法的基础上进行了一些改进,将CaCl2溶液换成TB溶液,将LB 培养基换成SOB++培养基,将摇菌温度改为18 °C,冻存时将15%甘油换成7%DMSO,可以更好地提高感受态细胞的效率。
我们实验室常用的感受态菌株主要有DH5α、BL21、Rosetta、DH10Bac,其中DH5α用于基因克隆,BL21、Rosetta用于原核表达,DH10Bac用于昆虫表达中Bacmid的制备。
下面以摇500 mL DH5α菌液,最终制备20 mL DH5α感受态细胞为例。
材料(MATERIALS)
r 试剂(REAGENTS)
无抗LB平板:20 g/L蛋白胨、5 g/L酵母提取物、5 g/L NaCl、15 g/L琼脂糖
5 mL LB培养基:20 g/L蛋白胨、5 g/L酵母提取物、5 g/L NaCl
DMSO
1 M MgCl2溶液(无菌)
1 M Mg MgSO4溶液(无菌)
500 mL SOB++培养基:20 g/L蛋白胨、5 g/L酵母提取物、0.5 g/L NaCl、0.186 g/L KCl、10 mM MgCl2、10 mM MgSO4(先将其他成分配好,高压蒸汽灭菌后再加入无菌的MgCl2和MgSO4)
120 mL TB溶液:10 mM HEPES(pH 6.7)、15 mM CaCl2、55 mM MnCl2、250 mM KCl(先将除MnCl2外的其他成分配置好,使用KOH将pH调至6.7,加入MnCl2后再使用0.22 μm的滤膜过滤除菌)
r 实验前准备(SETUP)
500 mL离心瓶
5 mL枪头
50 mL离心管
0.22 μm滤膜
一次性注射器
适量1.5 mL Ep管
r 器械(EQUIPMENT)
分液器、移液器
分析天平、pH计
超净工作台、摇床
Thermo落地式离心机(使用前预冷至4 °C)
实验步骤(PROCEDURE)
► 活化DH5α细胞 ►预计消耗时间:40 min(Day 1)
1.将超净工作台提前用紫外照射30 min,将无抗LB平板提前放入37 °C培养箱孵育。
2.从-80 °C冰箱迅速取出保藏的DH5α菌种,立即用一次性无菌接种环蘸取菌液,在孵育好的无抗平板上划线。
3.37 °C培养14~16 h。
► 物品准备,高压蒸汽灭菌►预计消耗时间:2 h(Day 2)
1.配置所需培养基和溶液,并将培养基、溶液和所需所有物品进行高压蒸汽灭菌。
2.预约后面一天的摇床,并提前对摇床进行紫外消毒。
► DH5α菌液小摇►预计消耗时间:40 min(Day 2)
1. 将超净工作台提前用紫外照射30 min。
2. 从平板上挑取DH5α单克隆菌落至5 mL无抗LB培养基中。
3. 放入37 °C摇床过夜摇菌。
► DH5α菌液大摇►预计消耗时间:2~3 h(Day 3)
1. 将超净工作台和37 °C摇床提前用紫外照射30 min。
2. 按照1:100将5 mL菌液加入到500 mL SOB++培养基中。
3. 18 °C,220 rpm摇菌,期间测量菌液OD600值,直至达到0.5左右。
▲ 关键步骤:
♫ 摇菌大约需要6 h。
♫ 18 °C时细菌生长更缓慢,形成的细胞壁更薄,更有利于吸收外源DNA。
♫ 37 °C时每20分钟,细菌增殖一代,OD600值翻倍,可作为参考,估算OD600值。
► 制备DH5α感受态细胞►预计消耗时间:2 h(Day 3)
1. 以下所有操作均在超净工作台中,且在冰上进行。
2. 将落地离心机4 °C预冷,将TB溶液放置于冰上预冷,将500 mL离心瓶、1.5 mL Ep管放入冰箱预冷。
3. 将菌液转移到500 mL离心瓶中,冰上放置10 min。
4. 4 °C,3000 g,10 min,离心,彻底弃去上清。
5. 用100 mL冰上预冷的TB溶液轻轻悬浮细胞,冰上放置30 min。
6. 4 °C,3000 g,10 min,离心,彻底弃去上清。
7. 用18.6 mL冰上预冷的TB溶液加入1.4 mL DMSO,轻轻悬浮细胞,冰上放置5 min。
8. 将1.5 mL Ep管依次插到冰上预冷。
9. 使用分液器以每份100 μL将感受态细胞分装到预冷的1.5 mL Ep管中,迅速扣上管子。
10. 迅速转移到液氮中冷冻,再转移到-80 °C中长期保存。
▲ 关键步骤:
♫ 操作要轻柔且迅速,插管和扣管子可以找人帮忙。
♫ 分装时,每次吸取菌液前都要轻轻摇匀。
► 效价检测►预计消耗时间:1.5 h(Day 3)
这里使用puc19质粒检测效价,将高浓度的质粒稀释到1 ng/μL,0.1 ng/μL。
污染检测 | ① | 分别将感受态涂布在Amp和Kan抗性平板上,37 °C过夜培养 |
② | 取少量感受态细胞加入到LB培养基中,过夜摇菌后涂布在无抗平板上,37℃培养,观察是否出现噬菌斑 | |
效价检测 | ③ | 将1 μL高浓度未稀释的puc19质粒转化到感受态后,加入900 μL无抗LB培养基复苏一个小时后吸取100 μL涂布在Amp抗性板上,37 °C过夜培养 |
④ | 将1 μL 1 ng/μL的puc19质粒转化到感受态后,加入900 μL无抗LB培养基复苏一个小时后吸取100 μL涂布在Amp抗性板上,37 °C过夜培养 | |
⑤ | 将1 μL 0.1 ng/μL的puc19质粒转化到感受态后,加入900 μL无抗LB培养基复苏一个小时后吸取100 μL涂布在Amp抗性板上,37 °C过夜培养 |
若①中未观察到菌落生长,②中未观察到噬菌斑,则说明未检测到污染。
转化子总数=菌落数×质粒稀释倍数×转化反应原液总体积/涂板菌液体积
计算感受态效率的公式为:⑤中长出来的克隆数×105。
一般⑤长出一百个以上克隆,那么感受态转化效率在7次方以上,单位为克隆数/微克。
针对性建议(TROUBLESHOOTING)
1. 所需实际可以提前配置好高浓度的母液。
2. 平板划线可以更好地活化细菌,最好不要省略这一步。
3. 当菌液OD600值接近0.5时,可以根据每隔20 min细菌增殖一代,OD值翻倍来估算一下,防止OD600值过高。
4. 分装时要迅速,分装后立即置于液氮中冷冻,再转移到-80 °C中长期保存。
5. 若感受态效价较高,可只检测①②⑤。
2.1.6 电转化感受态制备
简介(INTRODUCTION)
电转化(electrotransformation),也有人称作高压电穿孔法(high-voltage electroproration),简称电穿孔法(electroproration),可用于将DNA导入真核细胞(如动物细胞和植物细胞) 和原核细胞(转化大肠杆菌和其他细菌等)。电转化技术的主要优点在于:对于磷酸钙DNA共沉淀法以及其他技术难以转化的细胞,电穿孔法仍可适用。电转化的效率高,既可用于克隆化基因的瞬间表达,也可用于建立整合有外源基因的细胞系。本实验室曾使用电转化构建噬菌体展示文库,因此在电转化大肠杆菌方面积累了一定的经验,下面介绍的是电转化大肠杆菌的步骤。
材料(MATERIALS)
r 试剂(REAGENTS)
甘油,2xTY培养基(每1 L中含16 g蛋白胨、10 g酵母粉和5 g NaCl)
r 实验前准备(SETUP)
配置10%甘油,2xTY培养基,灭菌试剂及器械
r 器械(EQUIPMENT)
Thermo离心机、摇床、移液枪、锥形瓶、试管、离心瓶
实验步骤(PROCEDURE)
► 制备感受态
1. 挑取一个新鲜的需要制备感受态的菌株(例如TG1、DH5α等)克隆接种于3 mL 2×TY,37 °C、220 rpm培养6 h。
2. 1:100重新接种250 mL 2×TY,37 °C,200 rpm培养1 h 40 min至OD600为约0.6-0.8。
接下来的步骤均于冰面操作,所有的溶剂和仪器均需要提前预冷。
3. 将菌液转移至1 L离心瓶中,5000 g、4 °C离心15 min。
4. 倒掉上清,菌体用250 mL 10%的甘油重悬。5000 g、4 °C离心15 min。
5. 重复以上步骤一次。
6. 倒掉上清,菌体用100 mL 10%的甘油重悬。
7. 转移至2根预冷的50 mL离心管中。3000 g、4 °C离心10 min。
8. 倒掉上清,并彻底吸除残留的上清。
9. 加入1 mL 10%的甘油,用移液器轻轻吹打重悬。
10. 分装为100 μL每管(约12管),储存于-80 °C。
针对性建议(TROUBLESHOOTING)
制备过程中注意随时保持感受态处于低温状态。10%甘油需提前灭菌预冷。
2.1.7 氯化铷法感受态
简介(INTRODUCTION)
氯化铷(RbCl)法制备细菌感受态的转化效率被认为要高于常用的CaCl2法感受态,前者的转化效率可达到后者的10倍,RbCl法可能是目前效率最高的化学法感受态制备方法。除制备时所需的试剂配制较CaCl2法多一些外,RbCl法感受态的制备方法与CaCl2法大同小异。本实验室采用如下制备方法表明RbCl法感受态的转化效率为CaCl2法的2~8倍,本实验室使用的方案为金腾川教授提供。
材料(MATERIALS)
r 试剂(REAGENTS)
Buffer 1
成分 | 终浓度 | 总体积500 mL时加入量(g) | 总体积200 mL时加入量(g) | 总体积100 mL时加入量(g) |
RbCl | 100 mM | 6.046 | 2.42 | 1.21 |
MnCl2·4H2O | 50 mM | 4.95 | 1.98 | 0.99 |
KAc | 30 mM | 1.472 | 0.589 | 0.294 |
CaCl2·2H2O | 10 mM | 0.735 | 0.294 | 0.147 |
甘油 | 15 % | 75 mL | 30 mL | 15 mL |
水 | \ | 补足到500 mL | 补足到200 mL | 补足到100 mL |
加入约5~10 μl醋酸调节pH至5.8,0.22 μm滤膜过滤。
Buffer 2
成分 | 终浓度 | 总体积500 ml时加入量(g) | 总体积200 ml时加入量(g) | 总体积100 ml时加入量(g) |
MOPS | 10 mM | 1.046 | 0.419 | 0.209 |
RbCl | 10 mM | 0.605 | 0.242 | 0.121 |
CaCl2·2H2O | 75 mM | 5.513 | 2.205 | 1.103 |
甘油 | 15% | 75 mL | 30 mL | 15 mL |
水 | \ | 补足到500 mL | 补足到200 mL | 补足到100 mL |
加入HCl或NaOH调节pH至6.8,0.22 μm滤膜过滤。
SOC(500 mL)
成分 | 加入量 |
蛋白胨 | 10 g |
酵母粉 | 2.5 g |
NaCl | 0.25 g |
KCl | 0.1 g |
加ddH2O定容至200 mL,调节pH至7.0,高压灭菌20 min | |
2 M MgCl2(无菌) | 2.5 mL |
20% 葡糖糖(无菌) | 9 mL |
r 实验前准备(SETUP)
Buffer 1和2使用前应一直在放在4 °C冰箱预冷,准备灭菌的1.5 mL离心管,保证有足够的冰。
r 器械(EQUIPMENT)
离心机(使用前预冷至4 °C)。
实验步骤(PROCEDURE)
1. 挑取一个细菌单克隆接种3 mL LB,37 °C震荡培养过夜。
2. 1:100接种0.5 mL过夜菌至50 mL SOC培养基,37 °C震荡培养至OD600为0.5左右,一般需要1.5~2.5 h。
3. 转移菌液到离心管中,冰浴15 min,3000 g 4 °C离心10 min,倒掉上清。
4. 静置20 sec后吸走多余的上清。
5. 加入20 mL 预冷的Buffer 1,轻轻重悬细菌,冰浴30 min。
6. 3000 g、4 °C离心10 min,倒掉上清。
7. 静置20 sec后吸走多余的上清。
8. 加入4 mL预冷的Buffer 2,轻轻重悬细菌。冰浴15 min。
9. 冰面快速分装,每管100 μL。
10. -80 °C储存备用,1年内有效。
▲ 关键步骤:控制重新接种后的细菌生长,OD600不宜超过0.7。
针对性建议(TROUBLESHOOTING)
1.以上步骤中的细菌量可以按比例缩小或放大。
2. 制备好的感受态最好检测一下是否污染野生型噬菌体。
2.1.8 Trizol 法提取真核细胞总RNA
简介(INTRODUCTION)
利用传统的Trizol的萃取方法,可以从动物组织,真核细胞等中提取总RNA,进行RNA定量试验。样品被充分裂解后,加入氯仿离心后,溶液会形成上清层,中间层和有机层,RNA分布在上清层中,收集上清层经异丙醇沉淀后便可以回收得到总RNA。提取的总RNA纯度高,基本不含蛋白质和基因组DNA,可以用于mRNA纯化,体外翻译,RT-PCR等各种分子实验。
材料(MATERIALS)
r 试剂(REAGENTS)
DEPC水、氯仿、异丙醇、75%乙醇(DEPC水配制)、Trizol、PBS缓冲液
r 器材
RNase Free EP管和枪头、离心机、vortex、超净台、分光光度计、RNase Free研钵
实验步骤(PROCEDURE)
1.样品收集
1) 贴壁培养细胞:将培养基吸尽后,用4 °C预冷的PBS洗两遍后吸干净,加1 mL Trizol溶液吹打混匀用枪头将细胞刮下后,转移到1.5 mL离心管中,涡旋震荡30 s,冰上静置5 min充分裂解。
2) 悬浮生长细胞:细胞悬液倒入15 mL离心管中,800 g× 10 min离心,吸出培养基,加入2 mL 4 °C预冷PBS重悬细胞后再次800 g×10 min 4 °C离心,尽量吸干PBS,加入1 mL Trizol溶液吹打混匀,转移到1.5 mL离心管中,涡旋震荡30 s,冰上静置5 min充分裂解。
3) 组织样品:样品切成小块后用液氮速冻,在液氮预冷的研钵中充分研磨至粉末状,期间要不断加入液氮防样品融化。加入1 mL Trizol溶液混匀,继续研磨至裂解液透明,转移到1.5 mL离心管中,12000 g×5 min 4 °C离心吸取上清。
2.抽提RNA:
1) 加入1/5体积氯仿,剧烈振荡混匀30 s,使水相和有机相充分接触,冰上静置5 min。
2) 12000 g × 15 min 4 °C离心,可见分为三层,RNA在上层水相,中间层为DNA、下层为蛋白质。上层水相移至另一个新的RNase free EP管(吸上清时,枪头应沿着液面上层吸取上清,枪头不可碰到或吸到中间层,宁可少吸也不能吸到中间层)。
3) 沉淀RNA:加入等体积异丙醇,轻柔地充分混匀(颠倒6~8次,不应用振荡器混匀),冰上静置10 min。
4) 12000 g ×10 min 4 °C离心,去上清,收集RNA沉淀,沉淀为白色(如离心后仍不见EP管底部有沉淀,应将EP管放置在-80 °C冰箱过夜,继续在4 °C下,12000 g离心10 min,收集RNA沉淀)。
5) 用75%乙醇轻柔洗涤(加入乙醇后只需轻轻颠倒EP管即可,但要保证管壁、管盖都被洗涤到),轻弹管底使沉淀悬浮(不要用振荡器震荡或枪头吸打沉淀),冰上静置5 min。
6) 12000 g × 5 min 4 °C离心,尽可能去上清,室温超净台风干,沉淀会变为透明色,沉淀不能过干或过湿,过干则不易溶解,过湿则乙醇残留。
7) 视沉淀量加入适量DEPC水(至少15 μL)溶解沉淀,必要时可轻轻吹打。可-80 °C保存。
3. 总RNA纯度、浓度和完整性检测:
1) 纯度、浓度检测:取1 μL RNA样品,TE buffer或者DPEC水稀释,在核酸蛋白检测仪上测定OD值,OD260/OD280的比值大于1.8,说明制备的RNA较纯,无蛋白质污染。小于1.8则可能有DNA或蛋白污染。小于1.6可视为RNA为完全溶解。
2) 总RNA完整性检测:取RNA样品1 μL至8 μL TE buffer中,加DNA Loading buffer,1%琼脂糖凝胶电泳,用凝胶成像系统观察并拍照,如果是完整的RNA会有三个条带,分别是5 s rRNA、18 s rRNA和28 s rRNA。三条条带完整的话即可证明总RNA抽提比较完整。条带模糊或弥散则说明部分降解。
▲ 关键步骤:加入氯仿离心时切勿吸出白色中间层。RNA的抽提关键在于防止RNase污染降解RNA。

1.1.9 体外逆转录
简介(INTRODUCTION)
利用反转录试剂,可以在短时间内快速的将RNA高效合成用于Real-Time PCR用的cDNA。是进行Real-Time PCR反应的必须步骤。为了准确的进行RNA的定量表达分析,必须去除混杂的基因组DNA,该试剂盒利用gDNA wiper可以有效的去除基因组DNA,同时减少RNA的降解和损失。
材料(MATERIALS)
r 试剂(REAGENTS)
vazyme HiScript II Q RT SuperMix for qPCR(+gDNA wiper)Kit货号:R223-01
r 器械(EQUIPMENT)
PCR仪、PCR管、RNase Free枪头
实验步骤(PROCEDURE)
1. 去除基因组DNA。
2. 这里以vazyme HiScript II Q RT SuperMix for qPCR(+gDNA wiper)Kit为例操作:在RNase free的离心管中配制:
RNase free ddH2O | To 8 μL |
4× gDNA wiper Mix | 2 μL |
Oligo (dT)18 (10 μM) or Random hexamers (50 ng/μL) or Gene specific primers (2 μM) | 0.5 μL |
模板RNA | Total RNA:1 pg~500 ng |
轻轻吹打均匀,42 °C放置2 min。
3. 配制逆转录反应体系:8 μL上一步的反应液中直接加入2 μL 5×HiScript II qRT SuperMix II,轻轻吹打均匀。同时可选做不加逆转录酶的阴性对照反应(5×No RT Control Mix 2 μL),用于检测RNA模板中是否含有基因组DNA残留。
4. 在上一步的离心管中直接加入5×HiScript II qRT SuperMix II
5. 逆转录反应
25 °C | 10 min |
50 °C | 30 min |
85 °C | 5 min |
产物可立即用于PCR反应或-20 °C、-80 °C 保存半年,避免反复冻融。
▲关键步骤:
♫ 如果模板具有二级结构或高GC区域,可将第二部的反应温度提高到55 °C。
♫ 确保RNA是溶于水中而不是TE,因为TE对于基因组DNA的去除和逆转录的反应都产生抑制。
针对性建议(TROUBLESHOOTING)
试剂盒中的primer可以有效的反转录出cDNA,如果试验中需要用到特异性的反转录target,则需要单独进行specific primer的加样,不用重复加入试剂盒中的primer,同时将反转录的温度对应调节。
2.1.10 实时荧光定量PCR
简介(INTRODUCTION)
SYBR Green染料可以与双链DNA结合发出荧光,在实时定量PCR反应中,通过DNA链的热变性,引物退火,DNA聚合酶作用下引物的延伸三个步骤来扩增微量DNA,PCR仪器通过检测反应体系中的SYBR Green荧光强度,达到检测PCR产物扩增量的目的。
材料(MATERIALS)
r 试剂(REAGENTS)
vazyme SYBR Green Master Mix Kit 货号:Q111-02/03
r 器械(EQUIPMENT)
ABI Step one仪器(为例),八连管及管盖,离心机
实验步骤(PROCEDURE)
这里以vazyme SYBR Green Master Mix Kit为例
1.qPCR反应体系配制:
SYBR Green master Mix | 10 μL |
Primer1(10 μM) | 0.4 μL |
Primer2(10 μM) | 0.4 μL |
ROX Reference Dye 1 | 0.4 μL |
模板DNA | 2 μL |
ddH2O | To 20 μL |
2. RT-PCR反应:
两步法PCR扩增 |
|
Stage1: | 预变性,95 °C 30 s |
Stage2: | PCR反应 |
| 95 °C,5 s,60 °C,30 s。重复40个循环 |
3. 仪器使用
1) 提前10 min打开荧光定量PCR仪,电脑开机,打开软件Step One software v2.0。
2) 在开始界面的三列按钮中选择第一列的advanced setup。
3) 左侧Setup一栏里面点击Experiment properties选项进行修改:写入Experiment Name,选择stepone instument(48 wells),选择quantitation-comparative CT(△△CT),选择SYBR GREEN Reagents,选择Standard(~2 hours to complete a run)。
4) 点击Setup一栏中plate setup选项进行修改define targets & samples,然后选择assign targets and samples选项,通过选择,确定每孔的targets和samples的名称。
5) 然后点击Setup一栏中run method选项。
6) 检查无误后,在RUN一栏里点击start run按钮开始PCR。
4. PCR结束后将文件到处:界面左上file—export data—选择results以及其他选项,并选择存档位置,以.xls形式保存数据。
▲关键步骤:在操作八连管时,使用新的PE手套,再盖上八连管的盖子,因为仪器接受荧光信号通过管盖接收,不污染管盖可以保证误差。加入反应液盖好盖子后,确保无气泡,可在1500 g离心1 min。
2.1.11 基因定点突变
简介(INTRODUCTION)
定点突变(site-directed mutagenesis)是指通过聚合酶链式反应(PCR)等方法向目的DNA片段(可以是基因组,也可以是质粒)中引入所需变化,包括碱基的添加、删除、点突变(单点/多点)等。定点突变能迅速、高效的提高DNA所表达的目的蛋白的性状及表征,是基因研究工作中一种非常有用的手段。原理上分两种:1. 搭桥法(重叠PCR);2. 一步法(全质粒PCR)。
► 搭桥法(重叠PCR)定点突变
搭桥法共需要两对引物(两端引物,中间引物),三次PCR,其中前两次PCR可同时完成,原理如图一所示:两次PCR的产物回收,作为模板加上两端引物primer F和primer R 进行PCR3。
|
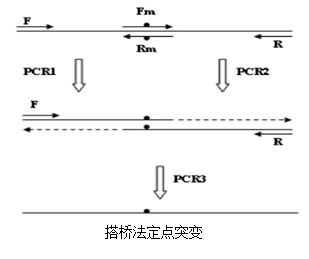
PCR1:以primer F和primer Rm为引物对扩增
PCR2:以primer R和primer Fm为引物对扩增
实验步骤(PROCEDURE)
1.两对引物的Tm值都应相当。两端PCR引物参照普通引物设计并无特殊要求。所需引入突变包含在中间引物互补区域内(需要在两条引物上均引入点突变),请勿将突变位点置于引物3’末端且突变位点距离3’端最少要有15个碱基,因为有非匹配碱基的存在,太短会导致引物与模板无法结合。
2. 对于一对中间引物的设计,如左图所示(高亮处是突变碱基),两引物间可以是完全互补,也可是部分互补。但两引物间互补部分的Tm值不能太低(太低导致PCR3无法配对延伸)。
5′-NNNNNNNNNNNNNNNANNNNNNNNNNNNNNNNN-3′ 3′-NNNNNNNNNNNNNNNTNNNNNNNNNNNNNNNNN-5′ 完全互补5′-NNNNNNANNNNNNNNNNNNNNNNN-3′ 3′-NNNNNNNNNNNNNNNTNNNNNNN-5′ 部分互补5′-NNNNNNANNNNNNNNNNNNNNNNN-3′ 3′-NNNNNNNNNNNNNNNT-5′ 部分互补 |
一对中间位置的点突变引物设计
3. PCR:PCR1:以primer F和primer Rm为引物对扩增;PCR2:以primer R和primer Fm为引物对扩增。两次PCR的产物回收,作为模板加上两端引物primer F和primer R进行PCR3。(注意前两次不能使用taq聚合酶,因为taq在产物3’端多加一个A,导致后续的PCR3出现移码突变)
4. 克隆:回收PCR3产物,酶切,连接,转化。
► 一步法定点突变
一步法是以质粒为模板,考虑扩增效率需将正向引物和反向引物分开扩增,避免二聚体的产生。原理如图。
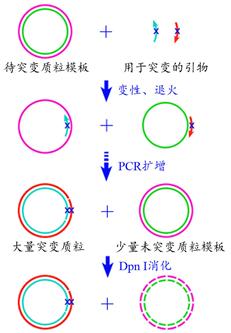
实验步骤(PROCEDURE)
1.引物设计:设计一对含有目标突变的引物,除所需要引入的突变位点外,其余序列与质粒模板完全匹配。不同于搭桥法的一对中间引物,全质粒PCR法的一对引物最好不是完全配对,因为这对引物是放在一个PCR反应中,完全配对极易形成引物二聚体,而不是与模板质粒结合。这就要求两条引物间配对区域的Tm值要小。
2.PCR:PCR1:质粒+正向引物扩增;PCR2:质粒+反向引物扩增。两次PCR均按照常规PCR程序进行,不同的是只反应12个而不是30个循环。PCR3:PCR1和PCR2反应完成后,将二者体系混合成一个体系并补加适量的高保真聚合酶(如Pfu或Pfu-X1),然后按照常规PCR程序继续反应16个循环。
▲关键步骤:
♫ 不能用taq酶,一是taq酶无3′-5′外切酶活性导致保真性差,全质粒PCR片段长,易出现非目标突变。二是因为taq酶具有5′-3′外切酶活性这就导致当PCR以质粒为模板复制一圈回到最初引物位置时,原有引物会被外切导致引入的突变又变回野生型质粒的序列。
♫ 为防止非目标突变,PCR循环数不要太多(建议不超过25个cycle)。
♫ 在质粒能正常扩增的前提下,应尽量减少质粒模板使用量以免模板质粒消化不完全背景增加。(酶切和PCR都无法区分)
3. PCR产物的甲基化酶处理
待突变的质粒通常来源于DH5α等dam+大肠杆菌,在这些dam+细菌中质粒会被甲基化修饰,而在体外通过PCR扩增得到的质粒不会被甲基化。这样用特异性识别甲基化位点的酶DpnI处理,可以消化掉待突变的质粒模板,而使通过PCR扩增出来的含有突变位点的质粒被选择性地保留下来(DpnI 酶只消化甲基化的DNA)。随后把DpnI处理过的产物转化感受态细菌后,突变质粒中的两个nick位点可以被大肠杆菌修复,得到的克隆就会含有预期的突变质粒了。
4. 向PCR产物中加入1 μL DpnI 酶和4 μL的10×NEB Cutsmart buffer消化模板质粒置于37 °C反应1 h,取出5 μL跑胶检测,剩余的取部分做细菌转化、涂板。筛选转化子、测序。
